

EVO DEVO

Les corps, et de surcroit leurs tissus biologiques sont intimement liés à leur environnement. Quelle que soit l’échelle considérée, nous sommes liés à celui-ci ; nos corps subissent la pesanteur, sont enveloppés dans l’atmosphère qu’ils respirent, de même ils agissent sur cet environnement en retour. Cette relation d’interdépendance n’étant pas unilatérale, nos comportements influent sur notre environnement proche et éloigné sans en avoir une conscience délibérée. Parfois de simples actions locales engendrent des phénomènes systémiques à une grande échelle nous affectant en retour.
Quel esprit suffisamment fécond peut s’imaginer qu’à tous les instants dans le monde macroscopique et microscopique, de la planète à la molécule, d’innombrables éléments vivants et non-vivants foisonnent à l’envi, interagissent directement et indirectement, créant, modifiant, multipliant, détruisant, des formes de vie diverses ?
Partout sur la planète, l’émergence de grâce faite d’harmonie, de légèreté et d'aisance dans les formes, dans la disposition et les proportions des parties, dans le mouvement, nous invite à l’élégance et à la beauté.
Les écosystèmes vivants, intriqués comme des poupées russes, interagissent à l’envi, de concert dans la création de formes de vie nouvelles, robustes et à la fois fragiles et ressemblant à s’y méprendre à ce qui a déjà été crée auparavant. Une fourmis née il y a des milliers d’années ressemble à celle de nos jours. C’est le paradoxe du vivant, l’alternance entre la plasticité et la stabilité des formes de la matière vivante.
Hormis aux échelles géologiques, comment parler d’évolution lorsque les formes de vies, de mêmes espèces, semblent identiques à l’échelle du vivant ?
Et à l’échelle du vivant, qu’est-ce qui crée les similarités et les différences entre deux embryons ?

Comment comprendre qu’il puisse exister des caractères communs, génétiques à un stade précoce embryonnaire pour ensuite créer une telle diversité de formes de vies ?
Tel un jeu de LegoTM la nature crée à partir de pièces identiques un assemblage différent et varié. Pour Jacques Monod (Monod 1970) : « la nature est objective et non projective. » en ce sens, elle « bricole » en dehors d’une conscience humaine et sans finalité, de façon hasardeuse, se soumettant pourtant à la nécessité des lois de la physique. Avant lui Démocrite disait : « Tout ce qui existe dans l'univers est le fruit du hasard et de la nécessité ».
Etudier l’évolution c’est à ce jour acter nos recherches dans une dynamique transdisciplinaire extrêmement féconde associant génétique du développement, embryologie, anatomie et paléontologie, ce que l’on nomme aujourd’hui « évo-dévo » pour évolution-développement.
Nous savons aujourd’hui qu’il existe une grande unité dans l’organisation du règne animal, et les recherches appellent à de nouveaux concepts sur l’origine des structures vivantes et sur la compréhension des transformations rapides se produisant au sein du vivant.
Irréversibilité des formes
Comment les tissus restent cohérents dans leur structure alors qu’ils font l’objet de remodelages constants, en profondeur, que les cellules se renouvellent et que les tissus vieillissent à la fois ?
Comment les formes s’établissent au cours du développement ? Quels sont les mécanismes génétiques, biochimiques, mécaniques qui gouvernent les assemblées cellulaires en développement ?
Comment fait un tissu pour associer le remodelage cellulaire constant (nos sommes des « ados cellulaires », (voir article) et l’irréversibilité des formes ?
Comment l’organisation et la dynamique d’un tissu à une échelle cellulaire fait pour créer l’émergence de formes complexes à une échelle supérieure ? Comment comprendre les interactions multiples mises en jeu entre les forces physiques, biochimiques participant au maintient des formes vivantes ?
Comment font les tissus pour conserver ordre et permanence de formes tout en favorisant leur évolution ?
Il est difficile de comprendre l'alliance paradoxale d'une organisation fixe et d'une dynamique incessante.

Formes et visages.
Tout le monde a reconnu : Clint, Meryl, Sigourney, Madonna, Jack, et Georges. Ils ont tous moins de 20 ans sur ces photos et ont dépassé la cinquantaine aujourd’hui et pourtant….
Comment les tissus restent cohérents dans leur structure alors qu’ils font l’objet de remodelages constant, en profondeur, que les cellules se renouvellent et que les tissus vieillissent à la fois ?
La ressemblance mère-fille est parfois aussi frappante. Vous reconnaissez aisément Catherine et Chiara, Demi et Scout Larue, Ingrid et Isabella, Tippi et Mélanie, Marlène et Eva, Lisa et Judy.

Formes et visages.
Dans la fratrie aussi.

Que je sache, aucun brave type ne se transforme en Brad Pitt spontanément sans passer par la chirurgie esthétique. La forme du visage, et sa reconnaissance de celui-ci par d’autres, c’est à dire notre représentation mentale liée à notre mémoire des visages (il y dans l’hippocampe des neurones dédiées à cela) reste constante et irréversible. Notre tronche, bouille, faciès, gueule, est notre identité et ceci pour longtemps.
Les visages semblent harmonieux n’est-ce pas ? Qu’en est-il réellement ? Si les tissus respectent une irréversibilité des formes facilitant la reconnaissance de ces visages, sont-ils pour autant symétriques ?

La symétrie des visages, des formes vivantes n’a rien de mathématique, ce qui n’empêche pas de garder des proportions parfois étranges qui « mathématiquement » correspondent à des critères de beauté.
La vie est une évolution. Le corps change de forme à tout instant. Ce qui est réel c'est le changement continuel de forme. La forme n'est qu'un instantané pris sur une transition.
Transformation des formes et formes en transformation
Une des propriétés du vivant est intrinsèquement dans la transformation, le mouvement, la dynamique, et l’évolution propre à toutes ces formes.
Les formes vivantes ont une structure changeante et à la fois une histoire propre. La pratique ostéopathique nous montre avec ferveur que la palpation fait émerger les formes particulières des corps, la spécificité de chacun, la rareté, la diversité mais aussi l’authenticité c’est à dire son histoire.
Lamarck en 1809 dans philosophie zoologique, a eu l’idée d’un cadre « physique » dans l’organisation du vivant. Les propriétés de la matière vivante proviennent de son organisation interne selon des lois physiques et du temps et que certaines fonctions, comme la pensée provient non pas de la matière elle-même mais de son agencement.
« Le génie souvent oublié de Lamarck fut de penser dans un cadre physique universel de propriétés essentielles du vivant à savoir : son organisation et son historicité. » (Thomas Lecuit 2017)
D’après de récentes recherches et selon T. Lecuit et V. Fleury (voir article), la forme de l’embryon et son développement représentent l’émergence d’une séquence de processus mécaniques reposant sur quatre notions : l’espace, le temps, l’information et la mécanique. L’image de l’origami convient à merveille, les plis, les forces, les opérations simples mais séquencés de façon précise tel un algorithme (une recette) donnent une forme majestueuse à partir d’un simple feuillet.

Une information de position définit les coordonnées cartésiennes de chaque cellule
Des amas de cellules regroupés selon leur taille au sein du disque embryonnaire, une information de lieu. (voir article)
Une information génétique définissant un espace et un temps donné, un ordre d’activation ou de répression des gènes présent dans la cellule (voir article) d’origine mécanique (mécanotransduction, mais pas seulement). Certains gènes dits homéotiques régulent en amont les processus de différentiation en aval. Ce sont des morphogènes (selon Alan Turing : une substance chimique productrice de forme). Ils traduisent l’information de position en information de différentiation. Ceci confère une orchestration génétique de la forme de l’embryon.
La forme émanant des interactions mécaniques loco-locales (mécanobiologiques) selon les principes d’intelligence collective (voir article) et d’auto-organisation (voir article) des assemblées cellulaires. L’orchestration se combine à l’auto-organisation dans les processus de développement.

Cathédrale sagrada familia et termitière cathédrale. Seuls les humains savent construire des cathédrales ?
L’information génétique organise, orchestre, dirige, ordonne l’activation ou la répression de certains gènes au sein de la cellule même. Cette information définit la position des cellules, les axes d’un embryon et le timing du développement des tissus. Mais il existe aussi une information mécanique, non génétique, provenant des interactions cellulaires locales (et avec leur environnement proche, notamment la rigidité de celui-ci) donc non orchestrée mais auto-organisée, qui fait émerger la forme du tissu.
« Qu'elle opère selon un principe d'orchestration à distance ou selon des règles locales d’auto organisation, l'information assure trois grandes fonctions :
- une fonction de régionalisation caractérisée par la définition de territoires par subdivision progressive,
- une fonction de polarisation par l'orientation de processus cellulaires dynamiques,
- une fonction de temporisation enfin par laquelle s'établit l'ordre temporel des processus. » (Thomas Lecuit 2017)
« Si l’information développementale est de nature chimique, n'oublions pas que les lois de la physique, en particulier celle de la mécanique, sont au cœur des interactions locales et à distance entre structures biologiques, de l'échelle moléculaire à l'échelle tissulaire et des organismes. » (Thomas Lecuit 2017)
Problème d’interface
Concernant les interactions loco-locales cellulaires à l’origine des formes des amas cellulaires et des tissus, il faut reconnaître l’apport des concepts mathématiques de D’Arcy Thomson (1860-1948), mathématicien écossais, biologiste, naturaliste, helléniste avec un don phénoménal pour les langues, pour qui les cellules utilisent des grandeurs physiques de tension de surface, d’énergie d’interface, avec leur environnement afin de créer des formes. Dans son livre On Growth and Forms publié en 1917, il postule l’idée selon laquelle les formes émergent d’un modèle thermodynamique lié à des transvections géométriques modifiant la forme des tissus au cours de l’évolution et ceci bien plus rapidement que selon les principes génétiques d’évolution Darwinienne.

De nombreux auteurs (Arkhat Abzarov) ont repris sa théorie, notamment sur les pinsons de Darwin, pour montrer que l’évolution et l’adaptation de la forme du bec des pinsons étaient bien plus rapides que les processus génétiques de mutations génétiques.
Depuis ces travaux, la biologie cellulaire a énormément évolué. L’imagerie a permis de visualiser à une résolution inégalée jusqu’à ce jour la dynamique du vivant. L’étude de l’embryon, et en particulier les épithéliums, connaissent des déformations majeures au cours du développement. Les forces adhérencielles maintiennent la cohésion cellulaire et tissulaire en lieu et place de ces déformations développementales.
Certaines molécules d’adhérence cellulaires, comme les E-cadhérines, sont responsables de cette cohésion. Plusieurs types de cadhérines existent au sein des tissus et sont classifiées selon ceux-ci. Les E-cadhérines : dans les cellules épithéliales et embryonnaires, les P-cadhérines : dans le placenta et l’épiderme, les N-cadhérines : dans les neurones et les muscles, les VE-cadhérines : dans le tissu vasculaire (endothélium). Elles ont un rôle dans les jonctions cellule-cellule et l’adhérence cellulaire.
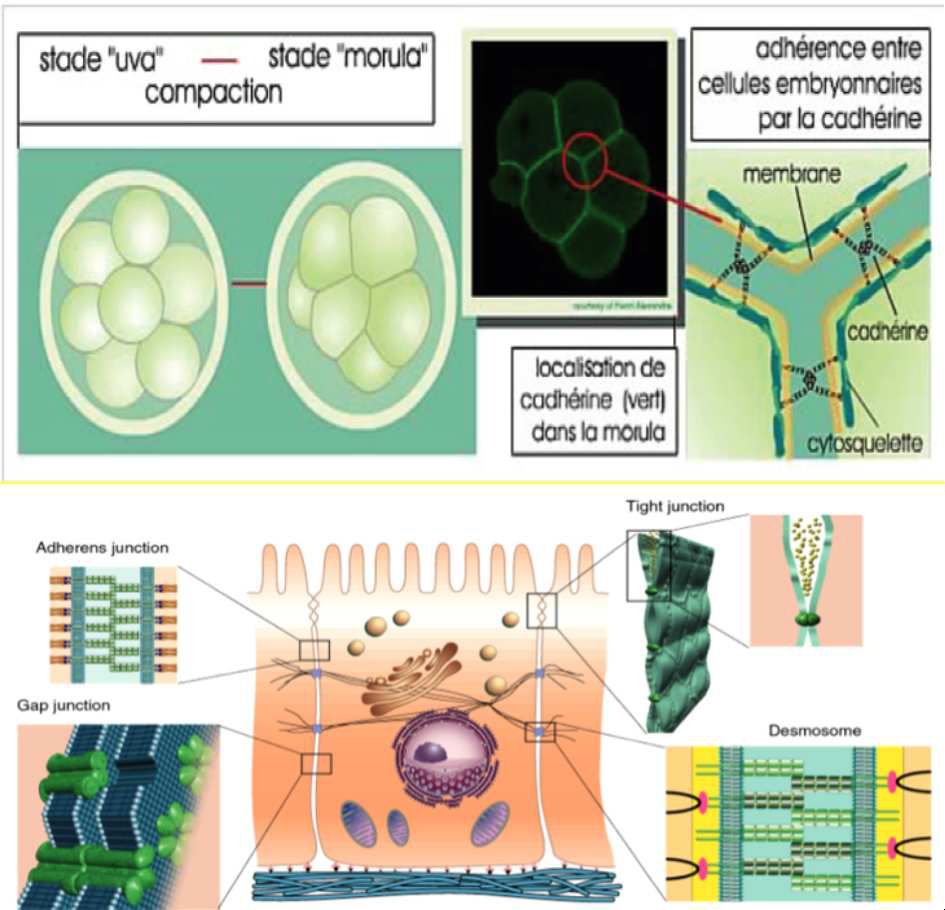

Ces molécules d’adhérence sont liées au cytosquelette cellulaire par l’intermédiaire des filaments d’actine ou intermédiaires selon le type de jonction cellule-cellule.
Dans les épithéliums particulièrement ces jonctions sont situées au niveau apical et latéral de ces amas linéaires monocouches.

« L’E-Cadhérine n'est pas distribuée de manière homogène et forme des agrégats, dont la taille et la localisation, la mobilité, sont régulées de façon locale. Le cytosquelette d'actine stabilise ces complexes et les immobilise. L'endocytose membranaire réduit leur taille, le réseau biochimique de polarisation cellulaire et les forces mécaniques ont des effets variables dont la caractérisation demeure encore incomplète. » (Thomas Lecuit 2017)
Etant reliées au cytosquelette, le dynamique de celui-ci interfère donc avec ces jonctions. Il existe un jeu dynamique entre la stabilisation des interfaces cellule-cellule et les forces contractiles du cytosquelette qui les déstabilisent.
C’est le paradoxe du vivant, l’alternance entre la plasticité et la stabilité des formes de la matière vivante.
Dans une structure épithéliale, les forces d’adhérence cellulaires, confèrent une géométrie particulière aux amas cellulaires, forme que l’on retrouve dans la plupart des structures à d’autres échelles, ananas, carapace de tortue, nid d’abeille, etc...

Remodelage et dynamique des interfaces
Le réseau d’actine du cytosquelette est rendu contractile à l’aide de moteurs moléculaires (les myosines II), qui se comportent un peu comme des muscles, car ce sont au sein de la fibre musculaire les mêmes éléments qui interagissent.
La cellule possède au sein de son cytoplasme (sorte de gel bourré de fibres) des ponts « acto-myosine » formant des dipôles responsables de la rigidité de celle-ci. On appelle cela des « fibres de stress », un peu comme des « petits muscles » plus ou moins tendus sur l’ensemble de la cellule. Elle se trouve ainsi précontrainte, c’est à dire ni complètement molle ni complètement rigide. Cette rigidité varie selon l’environnement dans lequel elle se trouve (voir article mécanobiologie).

Parfois ce réseau et ces ponts « acto-myosine » sont disposés plus près de la membrane (probablement grâce à d’autres molécules d’adhérence s’ajoutant à ces dipôles).
L’ensemble forme une coque d’environ 1 micron d’épaisseur entourant le cytoplasme, on parle de cortex d’actine. Dans certains mouvements cellulaires, comme l’extension d’un épiderme ou l’allongement d’un embryon, le taux de myosine II s’enrichit au niveau de ce cortex d’actine. (Rauzi. 2008)
Ainsi la forme de la cellule et ses jonctions avec les cellules voisines va changer. Dans le cas d’épithéliums à cellules longues et étroites, ce cortex est situé dans la région apico-latérale. De l’élaboration de l’architecture de la cellule émerge des déformations qu’elle subie autant venant de l’extérieur que de l’intérieur.
En fonction de l’axe de l’étirement, les « dipôles » formés s’alignent dans la direction de la traction provoquée. Ainsi tous les petits dipôles créent un ensemble de dipôles, comme un « macro-dipôle » conférant à la cellule, aux amas de cellules et aux tissus une polarisation.
En général les cellules s’alignent en fonction de la direction de l’étirement, mais ceci change ne fonction de la dynamique de cet étirement. Lors d’un étirement statique l’alignement se situe dans la direction de la contrainte appliquée au substrat. (Eastwood. 1998.)
Lors d’un étirement cyclique, l’alignement est perpendiculaire à la direction de la contrainte appliquée au substrat. (Yoshigi. 2005. Jungbauer. 2008)
Pour certains auteurs, les dipôles s’alignent dans la même direction que la sollicitation si celle-ci est relativement lente, si elle est trop rapide, les cellules s’orientent en direction perpendiculaire car elles n’ont pas le temps de s’aligner.
Pour d’autres auteurs, (Faust. 2011; De. 2007) l’alignement dépend de la fréquence de la contrainte appliquée. Les cellules s’aligneraient perpendiculairement à direction de la contrainte pour des fréquences supérieures à 1Hz.
L’adaptation à la rigidité du substrat, comme la traction cellulaire, est très courte, de l’ordre de1/10e de seconde quel que soit le niveau de modification de la rigidité.
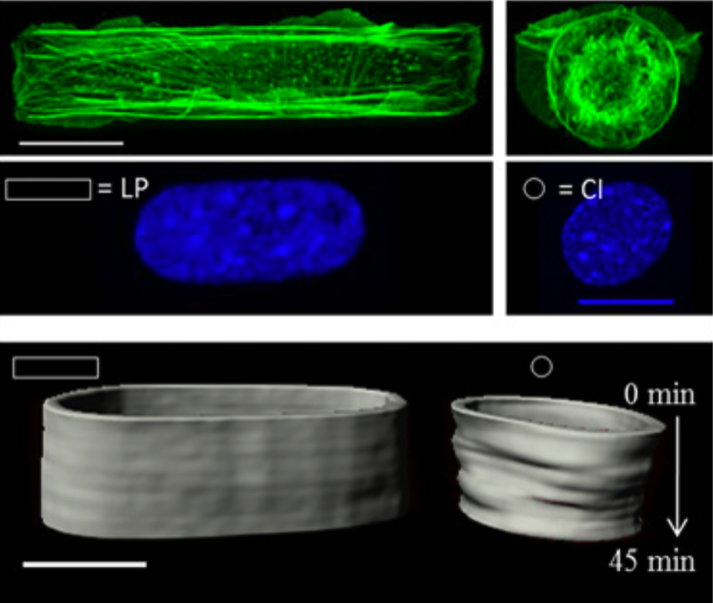
En cultivant des cellules sur des substrats de formes différentes, (Versaevel. 2012) en particulier allongées, le volume nucléaire est modifié affectant la compaction de la chromatine donc son expression.
Modification des interactions cellulaires au cours du développement
Dans l’extension d’un épithélium, les positions cellulaires suivent un processus d’intercalation. Elles perdent progressivement les contacts avec les cellules voisines dans le sens de l’élongation pour en créer avec celles qui sont situées perpendiculairement. On parle d’intercalation médio-latérale, on a ainsi un allongement du tissu qui devient plus étroit.
Un autre type d’intercalation existe lorsque le tissu s’affine en épaisseur et que des cellules de la profondeur viennent s’intégrer à celle de la superficie. On parle d’intercalation radiaire.
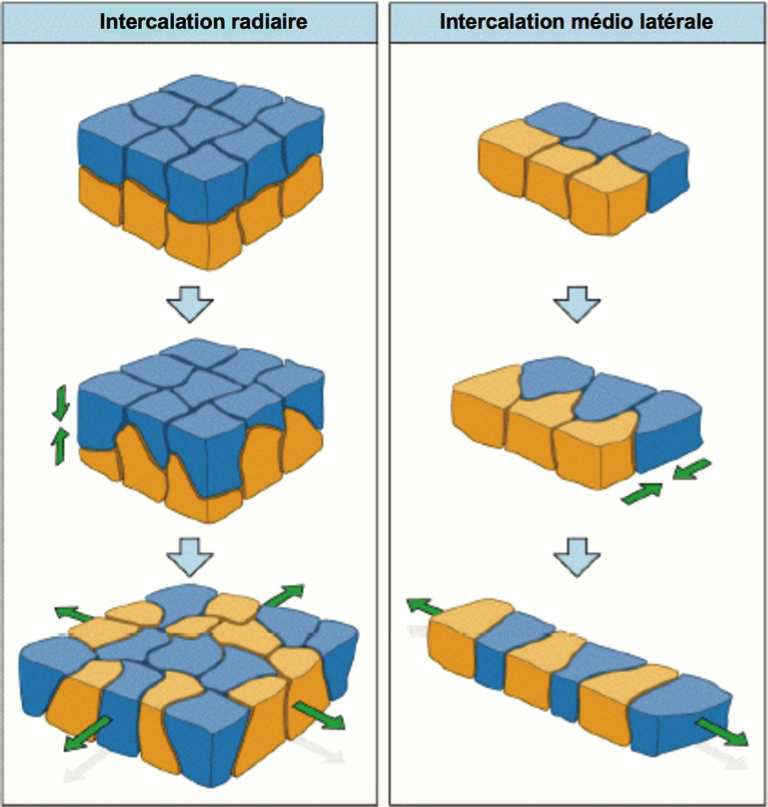

Exemple d’intercalation au cours de la gastrulation.
Dans l’embryogénèse, l’isotropie apicale induit une constriction et une invagination tissulaire, l’anisotropie induit une intercalation médio-latérale et un allongement tissulaire.

Guirao (Guirao 2015) a réussi à caractériser les processus cellulaires élémentaires dans la déformation des tissus.

D : division cellulaire, R : réarrangement, en fait cela correspond à une intercalation médio-latérale, S : size, grossissement cellulaire, A : apoptose, N : intercalation radiaire, C : fusion cellulaire, J : transformation en un autre type cellulaire
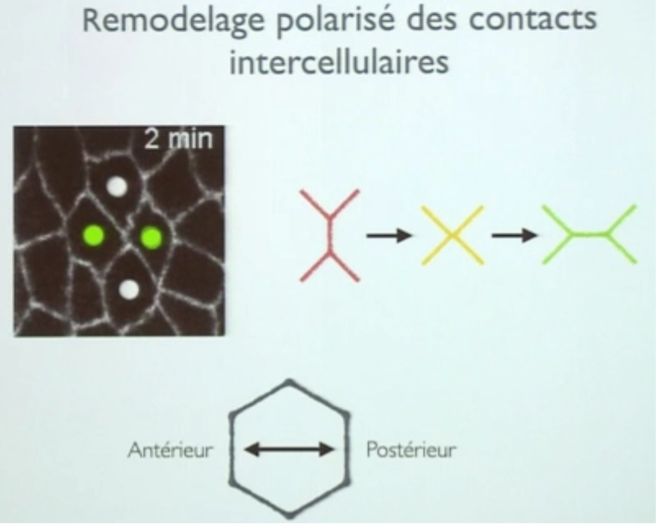
Ce remodelage et cette polarisation ne sont pas d’origine génétique essentiellement, mais dû aux interactions locales, donc il s’agit bien un processus auto-organisé. Il dépend de la modification des jonctions cellules-cellules liée à la tension mécanique provoquée par la proportion de myosine II locale et de la contractibilité du réseau d’acto-myosine.
D’autres auteurs (Makhijaa 2015) ont observé un accroissement de la dynamique de l’hétérochromatine et des télomères dans les cellules ou les sollicitations matrice-cellules sont réduites. Ainsi l’expression de la dynamique du cytosquelette, des lamines, modifie la dynamique chromatinienne.
Vous avez dit pulsations ?
La contractibilité du réseau d’acto-myosine n’a rien de fixe. Ces déformations sont pulsatiles, les contractions sont semi-périodiques. Lors de la formation d’une contraction, une infinité de moteur moléculaires et de filaments d’actine se regroupent en un endroit stratégique de la cellule, formant un flux migratoire local responsable de la future forme auto-organisée du tissu.
Les scientifiques parlent aujourd’hui de la notion de matière active, en ce sens, que ce processus auto-organisé se retrouve à toutes les échelles, des amas moléculaires en passant par les regroupements tissulaires jusqu’aux processus auto-organisés des bancs de poissons en pleine mer (voir intelligence collective).
Comme l’avait pressenti D’Arcy Thomson, la morphogénèse dépend aussi de paramètres physiques tel que l’adhérence, les forces contractiles comme la tension de surface corticale, la tension de surface intercellulaire et intertissulaire. Il en est de même pour l’organisation et la plasticité des tissus.
La morphogénèse c’est un large répertoire de comportements alternants déformation et stabilisation des formes.
« La rhéologie de la matière vivante est riche. Aux temps courts, quelques minutes, les cellules et les tissus sont des matériaux élastiques dans lesquels les déformations sont proportionnelles aux contraintes et réversibles, aux temps plus longs, on peut observer une gamme de comportements dissipatifs, par définition irréversible, reflétant la viscosité interne du matériau cellulaire et titulaire. » (Thomas Lecuit 2017)
Le secret de la matière vivante est à rechercher dans ces dynamiques déployées dans l’espace et le temps, à différentes échelles créant tissus et organes. C’est une alternance subtile entre des processus génétiques robustes et des processus auto-organisés d’influence mécano-biochimique et biologique créateurs de forme.
Finalement, biologistes et ostéopathes font le même travail. Pour reprendre une expression de Joël de Rosnay : « je cherche à comprendre » (De Rosnay 2017), que Jaques Monod a prononcé le jour de sa mort, d’un coté les biologistes observent et manipulent l’infiniment petit pour comprendre l’émergence des formes des tissus vivants à des échelles supérieures, et de l’autre coté les ostéopathes manipulent et observent à l’échelle humaine pour tenter de comprendre les propriétés immergentes de leurs actions sur les formes des tissus à des échelles inférieures. (Voir article les systèmes complexes)
Les propriétés émergentes apparaissent dans le sens « bottom-up », c’est à dire d’une échelle microscopique à une échelle macroscopique. L’association de différents éléments à caractéristiques particulières au niveau microscopique va constituer un ensemble à caractéristiques différentes. Dans l’autre sens, « top-down », du niveau macroscopique au niveau microscopique. Une action sur les systèmes en entier va créer une modification des propriétés des éléments constituants. Ce sont les propriétés immergentes.
Imaginez ce que pourrait donner le toucher ostéopathique, à la recherche d’un nouvel équilibre des tissus, d’une dynamique nouvelle sur le crâne ou tout autre partie du corps d’un nourrisson. Si, comme je le suppose, nos actions peuvent avoir un effet modifiant les propriétés immergentes en terme de qualité tissulaire, d’élasticité et de rigidité de façon plus harmonieuse, qu’en serait-il de l’évolution du bébé et de l’enfant en devenir ?
Nous sommes avec ces modèles mécanobiologiques bien au-delà de ce que certains, ostéopathes ou non, veulent bien nous enfermer. On ne parle plus de maladies, de douleurs, de techniques et de traitements, de restriction de mobilité, mais bien d’individualité, d’adaptabilité et de qualité.
Il faut bien comprendre que ces nouvelles approches en mécanobiologie donnent des modèles permettant de préciser la valeur des interactions mécaniques dans le biologique. Nous sommes au début d’une recherche probablement fructueuse, si les ostéopathes et les autorités sanitaires veulent bien s’en donner les moyens.
Je rêve d’un temps où les ostéopathes seront intégrés au sein des hôpitaux, en pédiatrie, en néonatologie, dans les services où ils pourraient étudier les effets de leurs actions sur des critères de jugement biochimiques ou biologiques. On pourrait ainsi objectiver nos actions, changer nos façons de faire, nos protocoles, en bonne intelligence avec les autres intervenants de ces services.
Références
Monod Jacques. Le hasard et la nécessité. Essai sur la philosophie naturelle de la biologie moderne. 1970.
Lecuit Thomas. Leçon inaugurale dynamiques du vivant. Collège de France. 2017.
Bergson Henri. 1907. L’évolution créatrice. Ed. Félix Alcan.
Lamarck JB. 1809. Philosophie zoologique : Exposition des considérations relatives à l’histoire naturelle des animaux. Dentu. Paris.
D’Arcy Thomson. On Growth and Forms. 1917
Rauzi, M., Verant, P., Lecuit, T., & Lenne, P.F. 2008. Nature and anisotropy of cortical forces orienting Drosophila tissue morphogenesis. Nature cell biology, 10(12), 1401– 1410.
Eastwood, M., Mudera, VC, McGrouther, DA, & Brown, RA. 1998. Effect of precise mechanical loading on fibroblast populated collagen lattices : morphological changes. Cell motility and the cytoskeleton, 40(1), 13–21.
Jungbauer, S., Gao, H., Spatz, J.P., & Kemkemer, R. 2008. Two characteristic regimes in frequency-dependent dynamic reorientation of fibroblasts on cyclically stretched substrates. Biophysical journal, 95(7), 3470–3478.
Yoshigi, M., Hoffman, L.M., Jensen, C.C., Yost, H.J., & Beckerle, M.C. 2005. Me- chanical force mobilizes zyxin from focal adhesions to actin filaments and regulates cytoskeletal reinforcement. The Journal of cell biology, 171(2), 209.
De, R., Zemel, A., & Safran, S.A. 2007. Dynamics of cell orientation. Nature Physics,
3(9), 655–659.
Faust, U., Hampe, N., Rubner, W., Kirchgeßner, N., Safran, S., Hoffmann, B., & Merkel, R. 2011. Cyclic Stress at mHz Frequencies Aligns Fibroblasts in Direction of Zero Strain. PloS one, 6(12), e28963.
Versaevel, M., Grevesse, T., & Gabriele, S. 2012. Spatial coordination between cell and nuclear shape within micropatterned endothelial cells. Nature Communications, 3, 671.
Boris Guirao. 2015. Unified quantitative characterization of epithelial tissue development. eLife. 4:e08519. DOI: 10.7554/eLife.08519
Ekta Makhijaa. D. S. Jokhuna. G. V. Shivashankar. 2015. Nuclear deformability and telomere dynamics are regulated by cell geometric constraints. PNAS. Dec 22.
De Rosnay Joël. 2016. Je cherche à comprendre. Les codes cachés de la nature. Les liens qui libèrent.



/image%2F1976184%2F20190501%2Fob_8cb8e7_sans-titre.png)
/image%2F1976184%2F20181117%2Fob_d7ffe2_sans-titre.png)
/image%2F1976184%2F20181016%2Fob_ad24b4_sans-titre.png)
/image%2F1976184%2F20180727%2Fob_c26024_sans-titre.png)
